正確なmRNAスプライシングを制御するU6 snRNAのm⁶A機構を解明 ――スプライシング異常が原因の疾患理解に貢献――
- ヘッドライン
- 記者発表
東京大学
発表のポイント
◆ 正確かつ効率的なmRNAスプライシングには、METTL16によるU6 snRNAのN6-メチルアデノシン(m6A)修飾が不可欠。
◆ METTL16、U6 snRNA、メチル基供与体SAMの三者複合体構造を解析し、METTL16のC末端KA-1ドメインがU6 snRNAを特異的に認識し、N末端の活性部位でのm6Aメチル化反応を促進する「足場」として機能することを発見。
◆ m6A修飾の欠失が引き起こすスプライシング異常に関連する疾患の分子基盤の解明に貢献。

METTL16・SAM・U6 snRNA三者複合体構造
概要
東京大学大学院新領域創成科学研究科メディカル情報生命専攻の琚 珏(ジュ ジュエ)特任助教と富田耕造教授は、前駆体mRNA(pre-mRNA)の正確かつ効率的なスプライシングに必須なU6 snRNAの特定アデニン(A)残基のN6位メチル化(m6A修飾)を担うメチル基転移酵素METTL16の反応分子機構を解明しました。U6 snRNAのm6A修飾は、pre-mRNAの5'スプライスサイト(5'-ss:mRNAのスプライシング開始点)のイントロン側配列と相互作用し、この相互作用を安定化することでスプライシングの正確性と効率性を確保します。
本研究では、METTL16のC末端KA-1ドメインがU6 snRNAの特定の領域を特異的に認識し、酵素にリクルートしてN末端の活性触媒部位でのメチル化反応を促進する詳細な分子機構が明らかになりました。この成果は、m6A修飾の欠失によるスプライシング異常に関連する疾患の分子基盤解明に貢献します。
本研究は、2025年8月21日付けで『Nature Communications』誌のオンライン版に掲載されました。
発表内容
N6-メチルアデノシン(m6A、注1)は、RNAに多く存在する修飾塩基で、RNAの細胞内局在、安定性、翻訳、pre-mRNAスプライシング(注2)など、遺伝子発現制御に重要な役割を果たします。mRNAのm6A修飾は主にMETTL3/METTL14複合体によって触媒されますが、METTL16はU6 snRNAなどの特定のRNA配列(ACAGAGAモチーフ:スプライシングに重要な7塩基のRNA配列、修飾アデニンAに下線)と構造を有するRNAを標的とするメチル基転移酵素です。U6 snRNAはpre-mRNAスプライシングに中心的な役割を果たす機能性RNAであり、U6 snRNA内の特定の位置のアデニン(A)のm6A修飾は正確なpre-mRNAスプライシングに必須です。m6A修飾の欠失は、U6 snRNAのACAGAGAモチーフと5'スプライスサイト(5'-ss:スプライシング部位)の相互作用を不安定化させ、スプライシングの効率と正確性を低下させ、イントロン保持を引き起こします。
METTL16は、ヒトから酵母まで保存されたメチル基転移酵素で、N末端のメチル化触媒活性ドメイン(MTD)とC末端のKA-1ドメインを有します。ヒトや線虫のMETTL16のKA-1ドメインは試験管内でU6 snRNAのm6A修飾を促進することが示唆されていましたが、その詳細な分子機構は未解明でした。本研究では、ヒトMETTL16と高い相同性を持つ分裂酵母Schizosaccharomyces pombeのMETTL16(spMETTL16)とU6 snRNAを用い、X線結晶構造解析、クライオ電子顕微鏡(cryo-EM、注3)解析、生化学的・機能的解析を行いました(図1、動画)。

図1:METTL16・SAM・U6 snRNA三者複合体構造
動画:https://drive.google.com/file/d/1dGZGfnkvyagILvBT7ME4E4GCakONRexX/view?usp=drive_link
(METTL16:MTD[マゼンタ]、KA-1[青]、SAM:シアン、U6 snRNA:オレンジ)
その結果、以下の(i)~(vi)が明らかになりました。
i) KA-1ドメインはU6 snRNAの内部ステムループ(ISL)領域に特異的に結合し、アルギニンに富む領域がISLの先端を特異的に認識します(図2a、2b、2c)。
ii) KA-1ドメインはU6 snRNAを酵素にリクルートする「足場」として機能し、m6A修飾活性およびpre-mRNAスプライシングに必須です。
iii) 生化学解析により、U6 snRNAが先にMETTL16のKA-1に結合し、その後メチル基供与体SAMがMTDの活性部位に結合することが判明。
iv) 初期結合段階:SAM非存在下では、U6 snRNAは伸長構造をとり、ACAGAGAモチーフ(メチル化されるAを含む)がMTDから離れた状態でU6 snRNAのISL領域を介してKA-1にリクルートされます(図2b、3a)。
v) 前生産段階(pre-productive stage):MTDへのSAMの結合によりMTDの構造が変化し、U6 snRNAのACAGAGAモチーフがMTDに認識され、活性触媒部位に近づきますが、メチル化されるAとSAMのメチル基は約6Å離れており、メチル化に至らない準備状態です(図2c、3b)。
vi) 生産段階(productive stage):U6 snRNAのテレステム形成やMETTL16のN末端伸長領域およびRNA結合ループとU6 snRNAの協調的相互作用により、ACAGAGAモチーフがSAMの結合したMTDの活性触媒ポケットに深く入り込み、効率的なm6A修飾が進行します(図3c)。
以上より、METTL16はKA-1ドメインを介してU6 snRNAのILSを特異的に認識し、U6 snRNAを酵素へリクルートし、その後のSAMのMTDへの結合による段階的な酵素およびU6 snRNAの構造変化を通じて m6A修飾を行うことが明らかになりました。

図2:METTL16・SAM・U6 snRNA三者複合体構造
(a) U6 snRNAの配列。ACAGAGAモチーフ(赤)内のA37がMETTL16によりメチル化される。構造解析で観察できなかった部分は灰色で示す。(b) SAM非存在時のMETTL16・U6 snRNA複合体(初期結合段階)。(c) SAM存在時のMETTL16・SAM・U6 snRNA複合体(前生産段階)。
(METTL16:MTD[マゼンタ]、KA-1[青]、SAM:シアン、U6 snRNA:ベージュ)
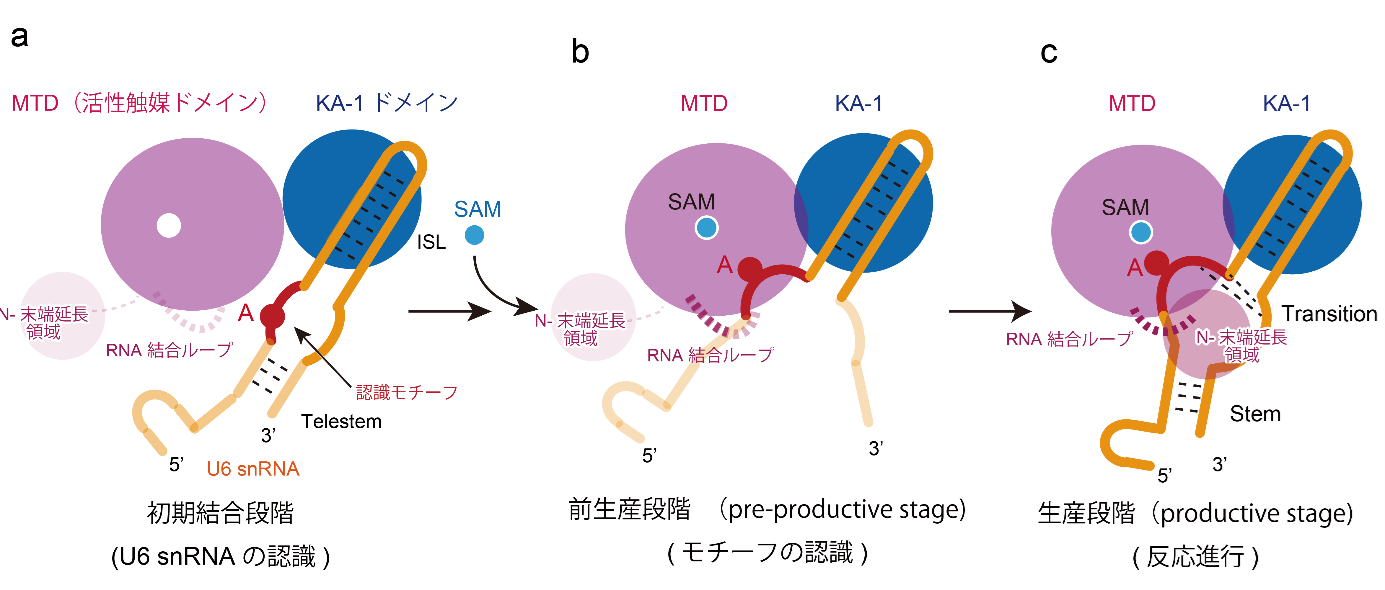
図3:METTL16によるU6 snRNAのm6A修飾の多段階反応
(a) 初期結合段階:U6 snRNAのISLがMETTL16のKA-1ドメインに認識される。(b) 前生産段階:SAMのMTD結合に伴いU6 snRNAのACAGAGAモチーフがMTDに認識される。(c) 生産段階:m6A修飾反応が進行する。
本研究は、METTL16がKA-1ドメインを介してU6 snRNAを特異的に認識し、多段階のタンパク質とRNAの構造変化を通じてm6A修飾を行う精密な分子機構を解明しました。KA-1ドメインは単なるRNA結合ドメインではなく、修飾活性の制御ハブとして機能します。このメカニズムは、KA-1ドメインがMETTL16のホモログ間で高度に保存されていることから、ヒトを含む真核生物で進化的・機能的に保存されていると考えられます。本成果は、RNA修飾による遺伝子発現制御の理解や、スプライシング異常に関連する疾患の分子基盤解明、新たな治療法開発に貢献します。
発表者・研究者等情報
東京大学大学院新領域創成科学研究科 メディカル情報生命専攻
琚 珏(ジュ ジュエ) 特任助教
富田 耕造 教授
論文情報
雑誌名:Nature Communications
題 名:Structures and mechanisms of U6 snRNA m6A modification by METTL16
著者名:Jue Ju and Kozo Tomita*
DOI: 10.1038/s41467-025-63021-0
URL: https://www.doi.org/10.1038/s41467-025-63021-0
研究助成
本研究は、日本学術振興会科学研究費助成事業、基盤研究A(課題番号:23H00368、18H03980)、文部科学省科学研究費助成事業 新学術領域研究(課題番号:26113002)国立研究開発法人日本医療研究開発機構(AMED)創薬等ライフサイエンス研究支援基盤事業 創薬等先端技術支援基盤プラットフォーム(BINDS)(課題番号:JP23ama121002)などの支援により実施されました。
用語解説
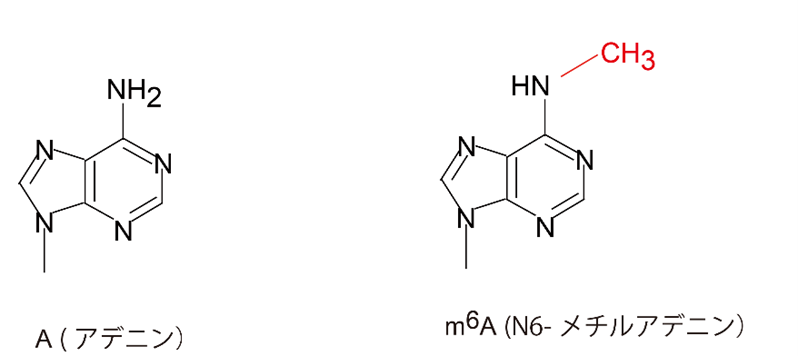
(注1)N6-メチルアデノシン(m6A)
RNAの塩基であるアデニンの6番目の窒素がメチル化されたヌクレオシド。mRNAの安定性、翻訳効率、スプライシングなどに影響を与え、遺伝子発現制御に重要。がんや神経疾患などさまざまな疾患に関連。
(注2)pre-mRNAスプライシング
真核生物において、DNAから転写された前駆体mRNA(pre-mRNA)から不要なイントロン(非コード領域)を除去し、エクソン(コード領域)を連結する過程。遺伝子発現の多様性や機能調節に寄与。
(注3)クライオ電子顕微鏡(cryo-EM)
試料を極低温で急速凍結し、電子顕微鏡で撮影した多数の画像から3次元構造を高分解能で再構築する技術。タンパク質や複合体の原子レベルの構造解析に用いられる。