◆受精後の最初の遺伝子発現は、これまで発生に寄与しないと考えられていたが、次の遺伝子発現を引き起こす役割を持ち、発生に必須であることを明らかにした。
◆受精後に起こる遺伝子発現の変化は、最初の発現が次の発現を引き起こすというステップ・バイ・ステップで調節されていることを明らかにした。
◆本研究により、受精後の遺伝子発現プログラムを調節するメカニズムの一端が明らかとなり、これが今後の全貌解明への端緒となることが期待される。
受精後の遺伝子発現プログラムの進行を調節するメカニズムの一端を解明
投稿日:2018/07/09
- ニュース
- 研究成果
- 記者発表
発表のポイント
発表概要
受精直後の胚では遺伝子発現が完全に停止していますが、一定時間を経過した後に最初の発現が起こります。その後、定まったプログラムに従い遺伝子発現パターンが変化して発生が進行して行きますが、このプログラムを調節するメカニズムについてはこれまでほとんど明らかにされていませんでした。
受精後に起こる最初の遺伝子発現(注1)は、まず小規模な胚性遺伝子の活性化(注2)と呼ばれる第1段階の転写活性化が起こり、次いで大規模な胚性遺伝子の活性化(注3)と呼ばれる第2段階の転写活性化と続くことにより進行します。そして、受精後にまず小規模な遺伝子発現の活性化が起こることは20年以上前に明らかにされていましたが、これまで発生には寄与していないものと考えられていました。しかし、今回、東京大学大学院新領域創成科学研究科の青木不学教授らのグループは、この発現を一過的に抑制することにより、それが次の大規模な遺伝子発現が起こすことに必要であり、さらに発生に必須であることを明らかにしました。この結果は同時に、受精直後の遺伝子発現プログラムがステップ・バイ・ステップで進行していることを示すものです。
以上、本研究により、これまでにほとんど明らかにされていなかった遺伝子発現プログラムを調節するメカニズムの一端が明らかとなり、これが今後の全貌解明への端緒となることが期待されます。
発表内容
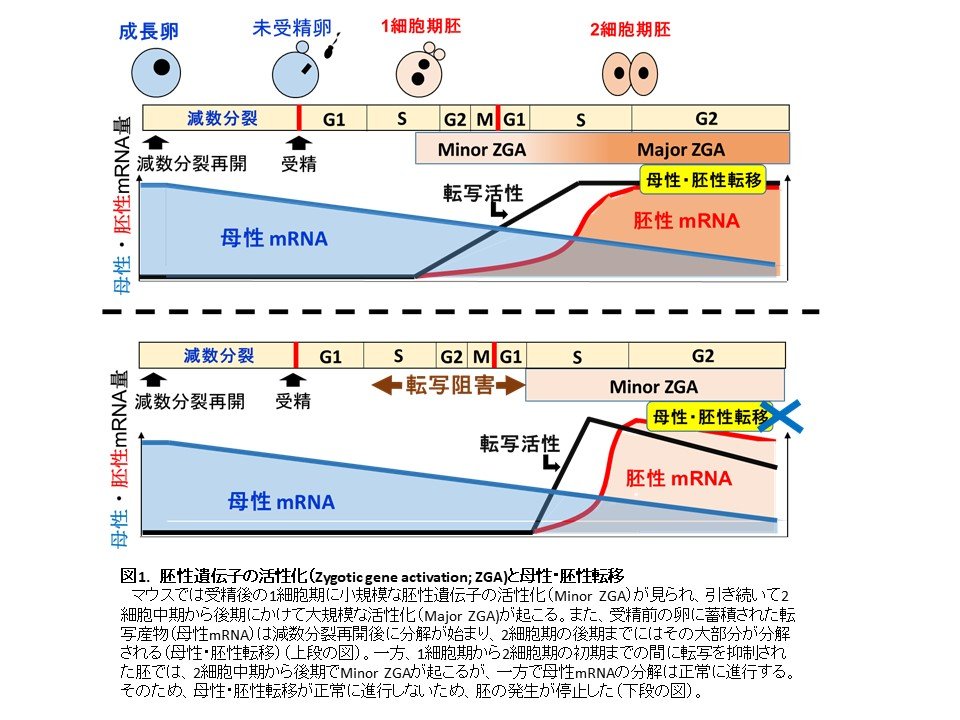
マウスでは、1細胞期から2細胞期のごく初期までに小規模な胚性遺伝子の活性化が起こり、大規模な活性化は2細胞期の中期から起こります。そこで、マウスの受精卵を用いて、1細胞期の転写を可逆的な試薬(5, 6-dichloro-1-β-D-ribofuranosyl-benzimidazole:DRB(注4))で一過的に抑制し、その後、2細胞期の初期に抑制を解除することで、受精後に最初に起こる転写の役割を調べました。その結果、DRBを含む培地からこれを含まない培地に戻された2細胞期胚は抑制が解除されて転写を開始しましたが、本来大規模な転写が見られる2細胞後期において、小規模な転写が起こることが明らかとなりました。また、1細胞期から2細胞期にかけてクロマチン構造の大規模な変化が起こりますが、DRBで一過的に転写を抑制された胚では、2細胞期でも1細胞期のようなクロマチン構造の特徴が見られました。一方で、受精前の卵から持ち越された転写産物(母性mRNA)は通常2細胞期の後期までにほとんどが分解されますが、DRBで一過的に転写抑制された胚でも同様にその分解が進んでいました。さらに、これらの胚はその大部分が2細胞期で発生を停止していました。これまでの知見では、2細胞期に起こる大規模な胚性遺伝子の活性化は発生に必要ですが、1細胞期の小規模な活性化は発生に関与していないと考えられていました。しかし、本研究により、大規模な遺伝子発現の活性化が起こる前に小規模な活性化が起こる必要があり、この小規模な活性化も発生に必須であることが明らかとなりました。
受精後の発生は、受精前の卵から持ち越された転写産物(母性mRNA)が分解され、それに胚生遺伝子の大規模な活性化により転写されたmRNAが置き換わることで調節されています(この現象は母性・胚性転移と呼ばれています)。しかし、一過的なDRB処理により、受精後最初の遺伝子発現が遅れて大規模な活性化が始まる前に母性mRNAの分解だけが進行し、この2つの時期にずれが生じたことで発生が進行しなかったものと考えられます(図1)。
また、受精後の遺伝子発現プログラムについては、その調節機構はほとんど明らかにされていませんでしたが、これまでプログラムの進行を規定するメカニズムとして次の3つが考えられていました。
(1)発生イベント:1細胞期から2細胞期、4細胞期と発生する段階、さらにはこれらのステージにおけるDNA合成、有糸分裂などのイベントによって遺伝子の発現パターンを変化させていくというものです。
(2)Zygotic Clock:発生イベントとは関係なく、受精後又はある発生時期からの経過時間が遺伝子発現プログラムの進行に関わっているという考えです。
(3)段階的変化:最初に発現した遺伝子産物が次の発現を引き起こし、これが順次続くことでプログラムが進行していくという概念です。このようなプログラムの進行がもっとも蓋然性があると考えられますが、実際には、この可能性を示す研究結果はこれまで報告されていませんでした。
ところが本研究において、1細胞期から2細胞初期まで一過的にDRBで処理したところ、発生後の時間は経過しており((2)zygotic clockは進行)、また母性mRNAの調節による発生も進行していました((1)DNA複製などの発生イベントも進行)。しかし、本来小規模な胚性遺伝子の活性化に続く大規模な活性化が見られる2細胞後期において、小規模な活性化が起こっており、大規模な活性化は見られませんでした。この結果により、遺伝子発現プログラムの開始時期においては、小規模から大規模な活性化へと段階的に遺伝子発現を変化させていくことが明らかとなりました。
以上より、これまでにほとんど明らかにされていなかった受精後の遺伝子発現プログラムを調節するメカニズムの一端が明らかとなり、これが今後の全貌解明への端緒となることが期待されます。
発表雑誌
雑誌名:Proceedings of the National Academy of Sciences, USA
論文タイトル:Minor zygotic gene activation is essential for mouse preimplantation development
著者:Ken-ichiro Abe, Satoshi Funaya, Machika Kawamura, Dai Tsukioka, Yutaka Suzuki, Masataka G. Suzuki, Richard M. Schultz and Fugaku Aoki
用語解説
東京大学大学院新領域創成科学研究科 教授 青木不学
TEL:04-7136-3695
Email:aokif@edu.k.u-tokyo.ac.jp
問い合わせ
(注1)受精後の最初の遺伝子発現
Zygotic gene activation (ZGA)と呼ばれている。
(注2)小規模な胚性遺伝子の活性化
Minor ZGAと呼ばれている。動物種によって起こる発生時期は異なっているが、多くの哺乳類では1細胞期に起こる。
(注3)大規模な胚性遺伝子の活性化
Major ZGAと呼ばれており、動物種によって起こる発生時期は異なっており、ヒトでは4~8細胞期、マウスでは2細胞期の後期に起こる。
(注4)5, 6-dichloro-1-β-D-ribofuranosyl-benzimidazole:DRB
遺伝子の転写の際に働くRNA polymerase IIの可逆的阻害剤。