紫外線を利用したアゲハ蝶の擬態術
- ニュース
- 記者発表
発表のポイント
◆毒蝶に擬態するシロオビアゲハのメスが、捕食者の鳥が認識できる紫外線(UV)を利用して、翅の白色紋様を毒蝶に似せるメカニズムを明らかにしました。
◆擬態を誘導するdsx遺伝子は、UV応答に関わる物質の合成だけでなく微細な鱗粉構造を調節し、擬態しないメスの白色領域(UVを吸収して青色蛍光を発する)を、UVを反射する擬態型の白色に変化させることがわかりました。
◆この発見は、UVの応答性を制御するような技術の開発の可能性や、アゲハ蝶の生活史や擬態の進化におけるUVの役割を示しました。
発表概要
東京大学大学院新領域創成科学研究科の依田真一特任研究員(研究当時)と藤原晴彦教授らの研究グループは、シロオビアゲハのメスが、紫外線(UV)を利用して毒蝶に巧みに擬態する分子機構を発見しました。
沖縄などに生息するシロオビアゲハには、翅の紋様を毒蝶のベニモンアゲハに似せた擬態型と、オスと似た紋様を示す非擬態型の2種類のメスがいます。擬態型メスと非擬態型メスの翅の中央部には白い斑紋がありますが、前者は毒蝶と同じようにUVを反射するのに対して、後者はUVを吸収して青く光ります。
擬態型メスで擬態を制御する遺伝子dsx(注1)の働きを抑制すると、UVを反射せずUVを吸収して青く光るようになりました。鳥などの捕食者はUVを見ることができることから、dsxはUVへの応答性を毒蝶に似せるように切替えていると考えられます。また、dsxはUV応答に関わる物質の合成だけでなく,微細な鱗粉構造も制御しています。この発見は、UVの応答性を制御するような技術の開発につながる可能性を示すとともに、アゲハ蝶の生活史や擬態の進化の過程でUVが大きな働きをしていることを明らかにしました。
これらの研究成果は、2021年1月8日付けでアメリカ科学振興協会AAASが発行するオープンアクセスジャーナル「Science Advances」にてオンライン公開されました。
発表内容
沖縄などに生息するシロオビアゲハには、毒蝶のベニモンアゲハに翅の紋様を似せた擬態型と、異なる紋様をもつ非擬態型の2種類のメスがいます。一方、オスは全て非擬態型のメスと類似した紋様を示します(図1)。150年近く前、進化生物学の先駆者ダーウィンとウォレスは、シロオビアゲハなどの大型の蝶ではメスだけが毒蝶に擬態する(ベイツ型擬態、注2)現象を不思議に思っていました。卵などを含むメスは栄養価が高く、動きも遅いため、鳥などの捕食圧が高く、オスに比べて擬態する利益が高いと言われていますが、メスだけが擬態できる分子的背景については最近までよくわかっていませんでした。シロオビアゲハの擬態を制御する遺伝子は100年以上前から、複数の遺伝子群が関与するsupergene(注3)からなることが指摘されていました。研究グループは、5年ほど前にゲノム配列や遺伝子機能解析を用いてこのsupergeneの構造を明らかにしました(注4)。
擬態型メスのシロオビアゲハの後翅の中央部には白い斑紋がある一方、非擬態型メスやオスでは白い帯状の紋様が見られます。私たちが見ると両者の白色はほとんど差がないように見えます。しかし、UVを照射すると、前者の白色はUVを反射するのに対し、後者の白色はUVを吸収して青い蛍光を発することが以前から知られていました(図2)。後者の蛍光色はパピリオクロームII(注5)という色素によるものと予想されました。そこで、エレクトロポレーション法(注6)を用いてパピリオクロームIIの合成経路の各遺伝子に対するsiRNA(注7)を非擬態型翅の白色部分で作用させて、RNAi(RNA干渉、注8)によって遺伝子発現を抑制しました。その結果,白色部分にUVを照射しても蛍光を発しなくなり、擬態型白色のようにUVを反射するようになったことから、非擬態型の翅の白色のUV応答性は主にパピリオクロームIIによって起こされると考えられました。
一方、擬態型を生み出すsupergeneの主要遺伝子であるdsxの発現を擬態型翅の白色領域で阻害すると、UVは反射しなくなり、UVを吸収して蛍光を発する非擬態型の翅に変化したこと(図3)から、擬態型翅ではdsxがパピリオクロームIIの生合成を抑制していることがわかりました。電子顕微鏡で調べると、擬態型の翅の鱗粉(注9)には隙間の多い骨格構造が見られるのに対し、非擬態型の翅の鱗粉にはパピリオクロームIIで埋められたような構造が見られます(図2)。dsxの発現を抑制するとこの構造も切り替わる(図3)ことから、dsxは物質の合成だけでなく鱗粉の物理的構造も制御していることがわかりました。興味深いことに、毒蝶のベニモンアゲハ後部の白色も擬態型メスの白色と同様にUVを反射し、その鱗粉も隙間の多い骨格構造が観察されました(図2)。隙間の多い骨格構造の鱗粉ではUVは反射され、パピリオクロームIIで充填された鱗粉だとUVは吸収されて蛍光を発すると考えられました(図4)。
私たちヒトはUVを見ることができませんが、捕食者である鳥やアゲハ蝶などはUVを受容する視細胞(注10)があり、UVを見ることができると言われています。従って、鳥や蝶から見ると、ベニモンアゲハの後翅の白色とシロオビアゲハの擬態型メスの後翅の白色はほとんど同じ「UV色」に見えるのに対し、UVを反射しない非擬態型の白色は全く異なる色に見えるはずです。図5はヒト、鳥、蝶からこれらの白色がどのように見えるかを推測してシミュレーションした結果を示したものです。3色しか見えないヒトでは3つの視細胞はそれぞれの白色に対して同じように応答しています。一方、鳥や蝶はUVを加えた4つの視細胞があり、特にUV視細胞での応答パターンは擬態型とベニモンアゲハの色に対してはよく似ているのに、非擬態型の色に対しては大きく異なります。このことは、UVが擬態の効果に寄与しており、鳥は毒蝶のベニモンアゲハにそっくりな擬態型のメスの捕食を避けようとすると考えられます。一方、シロオビアゲハのオスは自分と同じ模様の非擬態型メスを好むという説が以前からあり、両者がUVを吸収して青い蛍光を発するのはこの仮説を支持します。
今回の研究から、擬態の原因遺伝子dsxが物質の合成だけでなく電子顕微鏡レベルの微細な物理的構造を制御し、擬態型と非擬態型の翅のUVに対する応答を切替えていることがわかりました。これらの結果は、UVの吸収や反射に関して新たな物質や構造を創出するような応用に結びつく可能性があります。一方、UVが擬態のような種間コミュニケーションや性戦略といった種内コミュニケーションに大きな役割を果たしており、UVに対する応答性が遺伝的に制御され進化してきたことが明らかになりました。
発表雑誌
雑誌名:「Science Advances」(オンライン:2021年1月8日)
論文タイトル:Genetic switch in UV response of mimicry-related pale-yellow colors in Batesian mimic butterfly, Papilio polytes.
著者:Shinichi Yoda, Kousuke Sakakura, Tasuku Kitamura, Yusuke KonDo, Kazuki Sato, Ryosuke Ohnuki, Itsuki Someya, Shinya Komata, Tetsuya Kojima, Shinya Yoshioka and Haruhiko Fujiwara*
DOI番号:10.1126/sciadv.abd6475
発表者
依田 真一(研究当時:東京大学大学院新領域創成科学研究科先端生命科学専攻 特任研究員/現:基礎生物学研究所 特任助教)
藤原 晴彦(東京大学大学院新領域創成科学研究科先端生命科学専攻 教授)
用語解説
(注1)dsx遺伝子(doublesex):
脊椎動物から昆虫にいたるまで広く保存されている雌雄を分化させる遺伝子。下流の遺伝子の発現を制御する転写因子として知られる。シロオビアゲハには擬態を誘導するタイプの擬態型dsxと誘導しない非擬態型dsxが存在する。
(注2)ベイツ型擬態:
無毒な生物(擬態種)が系統的に離れた有毒な生物(モデル)に模様や形、行動などを似せて鳥などの捕食を免れる擬態。
(注3)supergene(超遺伝子):
擬態や社会性といった複雑な適応現象には、染色体の一カ所に集まる複数の遺伝子群で制御されている例が知られる。このような遺伝子群をsupergeneと呼ぶ。現在ではヒアリの社会性や鳥や魚の体表パターンなど数多くの現象がsupergeneにより制御されていることがわかっている。
(注4)Nishikawa et al. (2015) A genetic mechanism for female-limited Batesian mimicry in Papilio butterfly. Nat Genet. 47, 405-409. doi: 10.1038/ng.3241. において、シロオビアゲハのベイツ型擬態を制御する領域が3種類の遺伝子(dsx、U3X、UXT)を含むsupergeneで、dsx遺伝子が擬態を上位で誘導することを明らかにした。
(注5)パピリオクロームII:
アゲハ蝶の翅などで同定された色素物質。UV光で励起されて青い蛍光を発することが知られる。メラニン合成経路の産物のNBAD(N-beta-アラニルドーパミン)とキヌレニン合成経路の産物のキヌレニンが結合して形成される。
(注6)エレクトロポレーション法:
細胞や組織に極めて短い時間電気的な刺激を与えて細胞膜の一部に穴を開け、外部からDNAやRNAなどの物質を導入する方法。
(注7)siRNA:
RNAi(RNA干渉、注8参照)が起こる際に形成される21-21塩基ほどの短い二本鎖RNA。Small interfering RNAの略。siRNAは簡便に合成できるため、様々な生物種で特定の遺伝子機能を阻害するのに使われている。
(注8)RNAi(RNA干渉):
RNA interferenceの略。mRNAに相補的な塩基配列を持つ二本鎖RNAが、そのmRNAを分解する現象。遺伝子機能を阻害する方法の一つとして広く利用されている。
(注9)鱗粉:
蝶や蛾の翅は小さな鱗状の構造で覆われており、鱗粉と呼ばれる。それぞれの鱗粉は翅の表面の真皮細胞から分泌されたタンパク質やキチンなどが硬化して生じる。同時に色素が分泌されることもあり、これにより鱗粉の色が形成される。
(注10)視細胞:
光を受容する細胞の一種で、動物がものを見るときに光シグナルを神経情報に変換する役割をする。ヒトでは赤、緑、青に対する3種類の光受容細胞(それぞれL、M、S)があるが、鳥や蝶ではさらにUVを受容する細胞がある。
添付資料

図1:
シロオビアゲハ(Papilio polytes)のオス、非擬態型メス、擬態型メスと擬態型メスのモデルとなるベニモンアゲハ(Pachliopta aristolochiae)。
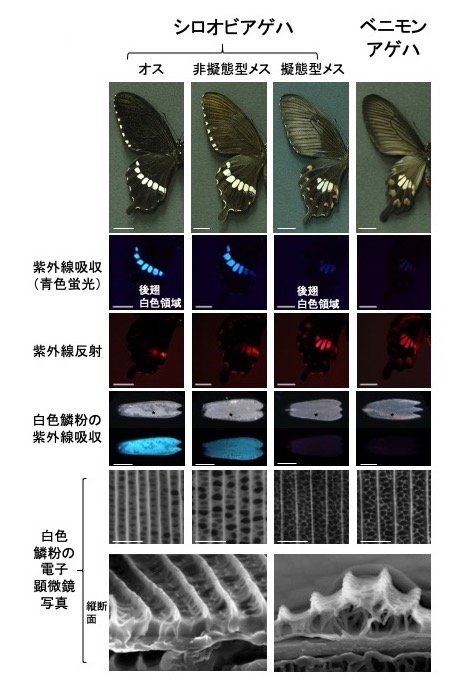
図2:
図1の4種の翅の紫外線応答の比較。上段から、翅のパターン、後翅白色領域の紫外線吸収(青色蛍光)、後翅白色領域の紫外線反射(赤色)、白色鱗粉の紫外線吸収(下段が紫外線照射時)、白色鱗粉の電子顕微鏡写真(下段は縦断面)。
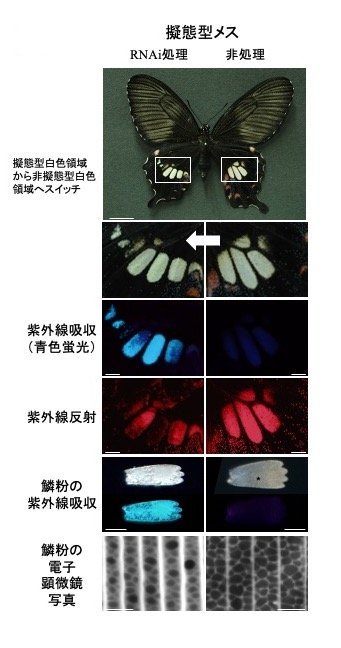
図3:
RNAi法によるdsxの発現抑制。擬態型dsx(注1参照)に対するsiRNAを蛹化直後の擬態型メスの蛹に注射後、エレクトロポレーション法で真皮細胞に導入した個体。左側が発現抑制した処理翅。右側は処理していないコントロール翅。発現抑制はsiRNAがモザイク的に導入されるため、効果の見られる部分と見られない部分が生じる。siRNAを導入した翅の白色領域の一部では紫外線を反射しなくなり、非擬態型メスと同様に紫外線を吸収して青色を発するように変化している。変化した鱗粉は紫外線で青色蛍光を発し、その電子顕微鏡写真は非擬態型と同様に見える。
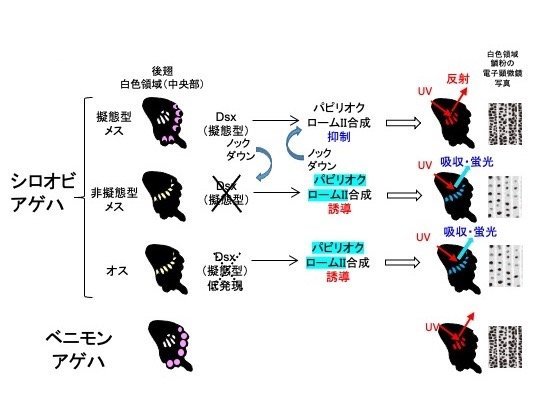
図4:
実験結果のまとめ。シロオビアゲハ擬態型メス(1段目)とベニモンアゲハの白色領域(4段目)は紫外線を反射し、鱗粉の電子顕微鏡構造は穴が多数空いたようにみえる。シロオビアゲハ非擬態型メス(2段目)とオスの白色領域(3段目)は紫外線を吸収して青色蛍光を発し、鱗粉の電子顕微鏡写真はパピリオクロームIIで埋まったように見える。擬態型メス白色領域ではdsx遺伝子はパピリオクロームIIの合成を抑制するが、その作用を阻害すると非擬態型メス型に変わる。非擬態型メス白色領域でパピリオクロームII関連遺伝子の発現を抑制すると擬態型メスと同様に紫外線を反射するようになる。オスではdsxの発現そのものが低く抑制されている。
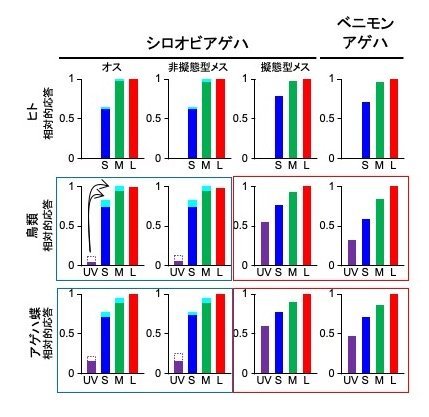
図5:
ヒト、鳥、アゲハ蝶の視細胞の応答。ヒトではL(赤)、M(緑)、S(青)の3種類の光受容体を持つ細胞があるが、鳥類とアゲハ蝶ではさらにUVを受容する細胞がある。シロオビアゲハオス、非擬態型メス、擬態型メス、ベニモンアゲハの白色領域の分光スペクトルを測定し、個々のデータをすでにわかっているシミュレーション数式(ヒト、鳥(アオガラ)、蝶(ナミアゲハ))に導入して、各細胞(横軸)の応答強度(縦軸)を相対的に算出した。ヒトでは4種類の白色の応答パターンはあまり変わらないが、鳥とアゲハ蝶では非擬態型メスとオス(青色線で囲む)、擬態型メスとベニモンアゲハ(赤色線で囲む)でパターンは類似している。UVを吸収して青色蛍光を発する変化は、それぞれ紫点線部と淡青色で表している(代表例として、図中に矢印で示した)。