地球全体に分布するロドプシン保有細菌の新たな光エネルギー獲得戦略 ― キサントフィルを用いた集光アンテナの発見 ―
- ヘッドライン
- 記者発表
東京大学
名古屋工業大学
発表のポイント
◆光エネルギーを化学エネルギーに変換するタンパク質(キサントロドプシン、プロテオロドプシン)が、カロテノイド色素の一種であるキサントフィル(ゼアキサンチンやルテイン)を集光アンテナとして利用することを発見しました。
◆ゼアキサンチンとキサントロドプシンとの複合体の立体構造を世界で初めて決定しました。
◆今回の成果から、集光アンテナを持つロドプシンが水圏微生物に広く分布し、ロドプシンによる光エネルギー受容量が従来の試算を大きく上回る可能性が示されました。
発表概要
地球上に存在するほぼ全ての生物は、太陽光由来のエネルギーを使うことで生命活動を行なっています。しかしながら、太陽光を生物が利用可能な化学エネルギーに変換できる生物は限られています。代表例は、クロロフィル色素を利用し光合成を行う生物(植物や一部の微生物)ですが、海洋や河川では光合成生物だけでなく、レチナール色素と結合した光受容タンパク質(微生物型ロドプシン:注1)を用いて太陽光を化学エネルギーに変換する微生物も数多く存在することが知られています。つまり、水圏環境ではこれら2種類の光受容機構が生態系に光エネルギーを取り込む窓口になっています。
今回、本研究グループは、イスラエル、スペイン、ドイツの研究グループとの国際共同研究により、淡水湖および海洋に生息する微生物が、レチナール色素に加えてカロテノイド色素(注2)の一種であるキサントフィル(ゼアキサンチンやルテイン)も結合するロドプシンを持つことを発見しました。さらに、これらの色素は受容した光エネルギーをレチナール色素に移動させる集光アンテナとして働くこと、集光アンテナを持つロドプシンが水圏微生物に幅広く分布することを明らかにしました。今回の発見により、水圏生態系においてロドプシンは集光アンテナを駆使し、従来の試算を上回る量の光エネルギーを受容することが示唆されました。生態系を理解する上で、光エネルギーを受け取る生物の光受容効率を把握し、生態系に流れ込む光エネルギー量を正確に算出することは避けては通れない課題です。本研究の成果は、全球レベルでの水圏生態系の理解の深化につながると期待されます。
本研究成果は、2023年3月1日(英国標準時)に英国科学誌「Nature」のオンライン版に掲載されました。
発表内容
【研究の背景】
ほぼ全ての生態系は太陽の光エネルギーを基盤に成立しており、生態系は光エネルギーを化学エネルギーに変換する生物によって支えられています。特に海洋や河川、湖沼などの水圏生態系では、植物プランクトンやシアノバクテリアなどの微生物が光合成を行なっており、これらの光合成生物のみが光エネルギーを変換できる、と長い間考えられていました。
ところが2000年、海洋細菌を対象としたメタゲノム解析(注3)から、光合成とは全く異なる、微生物型ロドプシンと呼ばれる光エネルギー受容機構の存在が報告されました。プロテオロドプシン(以降:PR)と名付けられたそのロドプシンは、結合したレチナール色素が光エネルギーを受容すると水素イオン(H+)を細胞の内側から外側へ輸送し、生物が利用可能な電気化学的ポテンシャルを形成します。その後の研究から、海洋表層に生息する細菌の過半数がPR遺伝子を保有し、特定の海域ではPRは光合成に匹敵するほどの光エネルギーを受容すると試算されています。また、微生物型ロドプシンは海洋だけでなく、河川や湖沼に生息する微生物からも次々と発見されており、生態系に流れ込む光エネルギー量を把握する上で、ロドプシンの光受容効率の理解は非常に重要だと考えられています。しかしながら、微生物型ロドプシンの光受容はレチナール色素のみが担うのか?それともレチナール色素にエネルギー伝達可能な集光アンテナを持つのか?などの、基本的な特徴すらよく分かっていませんでした。
【研究内容】
微生物型ロドプシンは一般的に、レチナール色素のみが光エネルギーを受容しますが、キサントロドプシン(以降:XR)は、レチナール色素の他にサリニキサンチンというカロテノイド色素と結合し、レチナール色素が受容できない青〜緑色の光を利用することができます。このような光受容を補助する色素は集光アンテナと呼ばれ、XRは一般的なロドプシンより幅広い波長域の光を受容できることが知られています。しかし、集光アンテナを備えたロドプシンは僅か3種類しか知られておらず、陸域の特定の環境に生息する微生物が持つ例外的な光受容機構であると考えられていました。また、既知の集光アンテナとして機能するカロテノイド色素は、ロドプシンタンパク質の立体構造上の制約から、全て4−ケト環構造(注4)という共通した構造を持ちます(図1)。
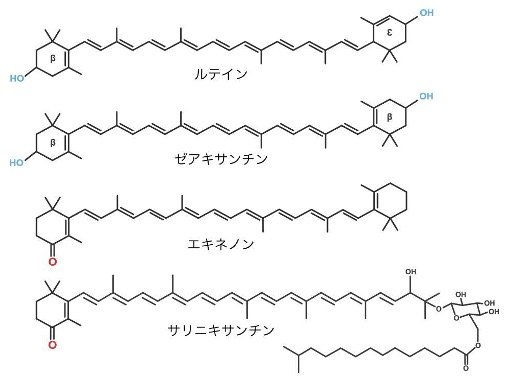
図1 カロテノイド色素の構造式
本研究により集光アンテナとして機能することが示されたゼアキサンチン、ルテインと、既知の集光アンテナであるサリニキサンチンとエキネノンの構造式の比較。ゼアキサンチンとルテインは4−ケト環構造(赤色の酸素原子)を持たない。
本研究では、陸域の特殊な環境以外に生息する微生物も、集光アンテナを備えたロドプシンを持つのかを調べました。イスラエルの淡水湖であるガリラヤ湖に生息する微生物を対象にメタゲノム解析を行い、ロドプシン遺伝子を探索しました。この遺伝子を大腸菌に異種発現(注5)させた後、精製したロドプシンタンパク質へ、湖水から抽出・濃縮した色素を添加しました。その結果、レチナール色素の他に、ゼアキサンチンやルテインと結合するXRを見出しました(図2)。さらに、このXRが利用できる光の波長を調べた結果、結合したゼアキサンチンやルテインが集光アンテナとして働き、受容した光エネルギーの約40%をレチナール色素に移動させることが明らかになりました。
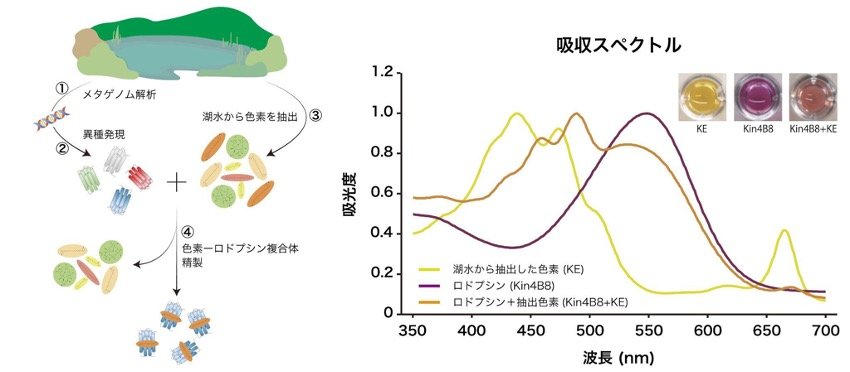
図2 実験の流れとカロテノイド色素と結合したキサントロドプシンの吸収スペクトル
左図は実験の流れ、右図は結果を示している。キサントロドプシンと湖水から抽出した色素の一部(ルテイン)が結合したことが分かる。
本研究で見出されたXRが、なぜ4−ケト環構造を持たないゼアキサンチンを集光アンテナとして使うことができるのかを調べるため、クライオ電子顕微鏡を用いた単粒子構造解析(注6)により、このXRの立体構造を決定しました(図3)。XRは5量体を形成しており、タンパク質の表面に大きな横穴が空いた単量体のそれぞれの外側に、ゼアキサンチンが結合していました。結合したゼアキサンチンの水酸基環は横穴にはまっており、レチナール色素と直接相互作用していました。また、ゼアキサンチンの水酸基は溶媒に露出しており、まわりの親水性アミノ酸であるセリンやチロシンと相互作用していました。一方、既知の4−ケト環構造を持つカロテノイドと結合するXRでは、これらの残基の極性は疎水性になっており、これらの横穴を構成するアミノ酸残基の性質や構造の違いが、XRの間で結合するカロテノイド色素の違いを決めていると考えられます。
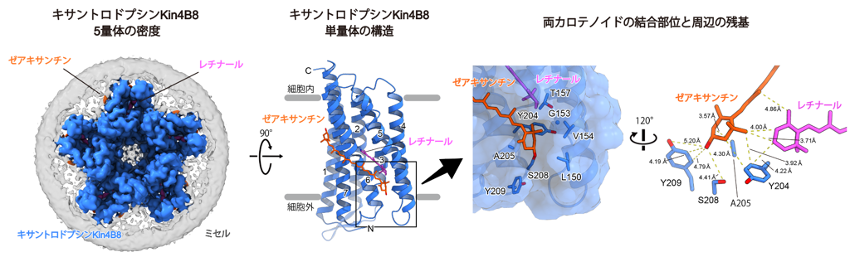
図3 ゼアキサンチンが結合したキサントロドプシンKin4B8の立体構造
左図は得られた5量体の密度マップと、モデル構築したKin4B8単量体の構造。右図はゼアキサンチンの結合様式を示している。
一方、地球表面積の約7割を占める海洋では、海洋表層に生息する細菌の半数以上がPRを保有することが知られていますが、その立体構造の特徴から集光アンテナを持たないと考えられていました。そこで学術研究船「淡青丸(注7)」を用いた研究航海で、西部北太平洋から分離した海洋細菌(Tenacibaculum sp. SG-28)を対象とし、SG-28株の持つPRが集光アンテナを備えるかどうかを調べました。SG-28株の持つPR遺伝子を大腸菌に異種発現させてタンパク質を精製した後、SG-28株から抽出したカロテノイド色素を添加し、再度タンパク質精製を行いました。一連の解析から、SG-28株が持つPRは、SG-28 株が産生するゼアキサンチンを集光アンテナとして用いることを示しました(図4)。
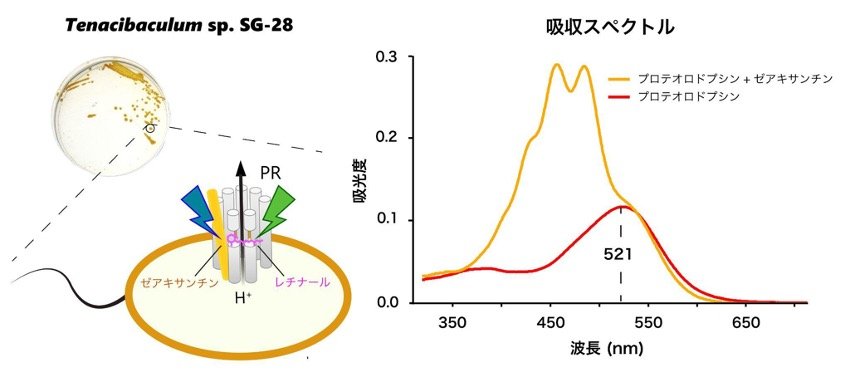
図4 集光アンテナによるプロテオロドプシン(PR)の光エネルギー受容量の変化
左図は海洋細菌Tenacibaculum sp. SG-28のコロニーの写真とSG-28株のもつプロテオロドプシンの模式図、右図はプロテオロドプシンの吸収スペクトルを示している。ゼアキサンチンと結合することで、プロテオロドプシンは短波長側の幅広い光エネルギーを受容することが出来る。
さらに、これまでに実施されたメタゲノムデータを利用し、各水圏環境から検出されたロドプシン全体に対し、集光アンテナを備える上で必要なアミノ酸を持つロドプシンの割合を調べました。その結果、集光アンテナを持つロドプシンは全世界の淡水域および、海域に広く分布することが明らかとなりました(図5)。
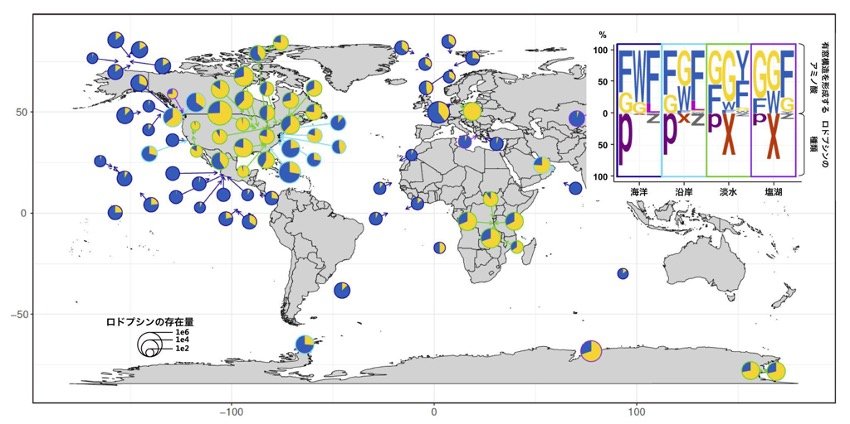
図5 集光アンテナを持つロドプシンの地理的分布
メタゲノムデータを用いて作成した、集光アンテナを備えたロドプシンの地理的分布を示している。ロドプシンが集光アンテナを備えるかどうかは、ロドプシンタンパク質上の特定の位置のアミノ酸の種類で推定することができる。この位置のアミノ酸がグリシン(G、黄色)であると横穴がある構造を形成し(=集光アンテナを備える)、それ以外のアミノ酸(フェニルアラニン(F)やトリプトファン(W)など、いずれも青色)であると横穴を形成せず、集光アンテナを持てない。ロドプシンのタイプはプロテオロドプシン(P、紫色)、キサントロドプシン(X、赤色)、それ以外のロドプシン(Z、灰色)を示す。海洋、沿岸、淡水、塩湖など幅広い環境に、横穴構造を形成するロドプシンが遍在すると推定される。
発表雑誌
雑誌名:「Nature」(オンライン版:3月1日)
論文タイトル:Phototrophy by antenna-containing rhodopsin pumps in aquatic environments
著者:Ariel Chazan#, Ishita Das#, Takayoshi Fujiwara#, Shunya Murakoshi#, Andrey Rozenberg#, Ana Molina-Márquez, Fumiya K. Sano, Tatsuki Tanaka, Patricia Gómez-Villegas, Shirley Larom, Alina Pushkarev, Partha Malakar, Masumi Hasegawa, Yuya Tsukamoto, Tomohiro Ishizuka, Masae Konno, Takashi Nagata, Yosuke Mizuno, Kota Katayama, Rei Abe-Yoshizumi, Sanford Ruhman, Keiichi Inoue, Hideki Kandori, Rosa M. León, Wataru Shihoya*, Susumu Yoshizawa*, Mordechai Sheves*, Osamu Nureki*, and Oded Béjà*
#(共同)筆頭著者、*責任著者
DOI番号:10.1038/s41586-023-05774-6
アブストラクトURL:https://www.nature.com/articles/s41586-023-05774-6
発表者
藤原 敬允(東京大学大学院新領域創成科学研究科 博士課程)
村越 峻也(東京大学大学院理学系研究科 修士課程)
井上 圭一(東京大学物性研究所 准教授)
神取 秀樹(名古屋工業大学大学院工学研究科 特別教授)
志甫谷 渉(東京大学大学院理学系研究科 助教)
吉澤 晋(東京大学大学院新領域創成科学研究科/東京大学大気海洋研究所 准教授)
濡木 理(東京大学大学院理学系研究科 教授)
用語解説
(注1)微生物型ロドプシン
7回膜貫通型の光受容タンパク質であり、発色団としてレチナール色素と結合する。ロドプシン遺伝子は細菌、古細菌、真核微生物、巨大ウイルスに広く分布することが知られている。
(注2)カロテノイド色素
主に400~500 nmの波長の光を吸収し、黄色、橙色、赤色を示す天然色素群。
(注3)メタゲノム解析
海水などの環境サンプルから抽出したDNAの塩基配列を網羅的に解読し、培養実験を介さずにサンプル中の微生物を解析する方法。解読されたDNAの配列データはメタゲノムデータと呼ばれる。
(注4)4−ケト環構造
炭素が形成する6員環構造において、4番目の炭素原子がケトン基をもつ構造。これまでに立体構造が決定された集光アンテナを持つロドプシンでは、カロテノイド色素のケトン基がロドプシンとの結合部位を形成している。
(注5)異種発現
目的遺伝子のDNAを人工的に他の生物に組み込んで、特定のタンパク質を作らせる実験手法。
(注6)単粒子構造解析
電子顕微鏡で撮影した多数の生体分子の画像から、その立体構造を決定する構造解析手法。目的試料の結晶を作製しなくても立体構造情報を得ることができる。
(注7)淡青丸
独立行政法人海洋研究開発機構が所有していた学術研究船。本研究で用いた海洋細菌(Tenacibaculum sp. SG-28)はKT-09-11次研究航海で分離し、研究室で維持してきたサンプルである。